
Membrana Plasmática
Tema 1
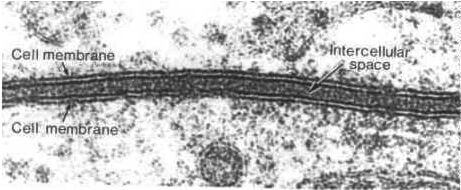
Ultra estrutura da membrana plasmática

A membrana plasmática / plasmela / membrana celular é a estrutura que envolve as células de todos os seres vivos. Delimita o meio externo e o meio interno e mantém a integridade celular.
A membrana plasmática não é totalmente impermeável. É como uma barreira seletiva, onde ocorrem trocas entre a célula e o meio exterior, permitindo que algumas substâncias como o oxigénio e nutrientes entrem, e outras, como os resíduos, saiam.
A membrana plasmática também funciona como um sensor, o que permite à célula modificar-se como resposta a estímulos ambientais.
A sua espessura (6 a 10 nm) é inferior ao poder de resolução do microscópio ótico, ainda assim já se sabia da existência da membrana plasmática antes da invenção do microscópio eletrónico. Por exemplo, A variação de células colocadas em diferentes concentrações apontava para a existência de um invólucro com uma certa plasticidade.
Constituição da membrana plasmática
O isolamento de membranas plasmáticas através de técnicas especiais permitiu identificar os seus constituintes. As membranas plasmáticas são complexos lipoproteícos, constituídos por proteínas (60% a 75%), lípidos (25% a 40%) e glícidos (podem chegar até aos 10%). Esta composição varia de célula para célula, relativamente ao peso.
Proteínas - Têm várias composições e funções. Estas moléculas podem ter funções estruturais ou interferir no transporte de substâncias na membrana e funcionarem como recetores de estímulos químicos do meio extracelular, ou como enzimas, estimulando reações que ocorrem na superfície da membrana plasmática.
Glícidos - Encontram-se na parte externa da membrana. Apesar de não se saber muito sobre as suas funções, são importantes no reconhecimento de algumas substâncias por parte da célula.
Lípidos- Maioritariamente fosfolípidos, colesterol e glicolípidos (lípidos associados a glícidos), em menor quantidade.
Ambos os fosfolípidos e os glicolípidos são moléculas anfipáticas, isto é, são constituídos por uma parte hidrofílica (polar) - tem afinidade com as moléculas da água, e por uma parte hidrofóbica (apolar) - não tem afinidade com as moléculas da água.
O colesterol faz parte do grupo dos esteróides - lípidos com uma estrutura contendo anéis de carbono. Insolúvel em água.

Modelos de estrutura da membrana plasmática
Apesar de se saber a composição molecular da membrana celular, não se sabe tanto sobre a sua organização estrutural, por ser bastante complexa. Assim, desde o século XX que vários modelos de estrutura da membrana celular têm sido apresentados e substituídos por outros, à medida que os conhecimentos sobre a estrutura, funções e composição membranares vão aumentando.
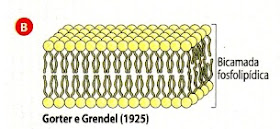
Modelo de Gorter e Grendel
Estudos feitos mais tarde com eritrócitos mostraram que a quantidade de fosfolípidos que estavam isolados da membrana era suficiente para formar uma dupla camada na superfície de cada eritrócito.
Estes estudos levaram Gorter e Grendel, em 1925, a apresentar um modelo de organização estrutural da membrana plasmática, baseado numa camada fosfolipídica. Os cientistas propunham que a membrana era composta por duas camadas de fosfolípidos onde as extremidades hidrofóbicas (apolares) ficariam viradas para o interior da membrana e as extremidades hidrofílicas (polares) ficariam voltadas para o exterior da membrana, havendo assim contacto com o meio externo e interno da célula.
Primeiro Modelo de Davson e Danielli
O modelo proposto por Gorter e Grendel foi mais tarde revisto por Davson e Danielli. Evidencias a respeito da permeabilidade e tensão superficial da membrana levaram a crer que as membranas seriam estruturas mais complexas que apenas camadas lipídicas. Em 1935, Davson e Danielli propuseram um modelo onde a bicamada fosfolipídica seria coberta interna e externamente por uma camada proteica que se ligaria às extremidades hidrofílicas dos fosfolípidos.
.PNG)
Estrutura da membrana ao microscópio eletrónico
Com o aparecimento do microscópio eletrónico, surgiram nos anos 50 as primeiras microfotografias de membranas obtidas através da técnica de fixação de tetróxido de ósmio e ampliadas mais de 100 000 vezes. Estas microfotografias pareciam confirmar o modelo proposto por Davson e Danielli, apresentando uma estrutura formada por duas linhas escuras separadas por uma banda clara.

As linhas escuras corresponderiam às proteínas e ás partes hidrofílicas dos fosfolipídos, e a banda clara corresponderia às partes hidrofóbicas dos fosfolipídos.
Ainda assim, este modelo não explicava a passagem de moléculas polares através da membrana, como a água, os aminoácidos e os monossacarídeos, pois uma camada contínua de fosfolipídos não seria atravessada por essas substâncias.
Segundo Modelo de Davson e Danielli

Em 1954, Davson e Danielli alteraram o modelo
inicial. A membrana celular teria poros revestidos por proteínas que formariam
passagens hidrofílicas por onde as substâncias
polares atravessavam e as substâncias não polares pela atravessavam a membrana
diretamente através da bicada fosfolipídica.
Como a pesquisa sobre a membrana plasmática não parou, alguns dados contribuíram para que o modelo proposto pelos dois cientistas fosse posto em causa:
- Análises quantitativas mostraram que as proteínas não seriam suficientes para cobrir toda a bicamada fosfolipídica.
- Averiguou-se que quando a membrana era sujeita a ações enzimáticas, a camada fosfolipídica era mais facilmente danificada que as proteínas;
- Verificou-se que algumas proteínas de destacavam facilmente da membrana, enquanto outras eram dificilmente removidas;
- Confirmou-se que as proteínas da membrana tinham regiões hidrofílicas e hidrofóbicas. Se as proteínas se encontrassem na superfície dos fosfolipídos, isso faria com que algumas regiões hidrofóbicas estivessem em contacto com a água.
Modelo do Mosaico Fluido
Todos estes dados levaram à proposta, em 1972, de um modelo da membrana, sendo o mais aceite hoje em dia. Este modelo foi proposto por Singer e Nicholson e designa-se Modelo do Mosaico Fluido.
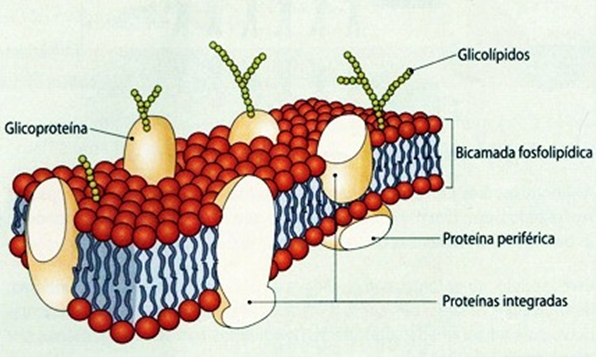
Este modelo admite que a membrana não é uma estrutura rígida, mas sim de grande fluidez, devido á existência de movimentos por parte das moléculas que a constituem.
Os fosfolipídos não estão fixos nas camadas, pelo que detêm grande mobilidade lateral, trocando de posição com outros fosfolipídos da mesma camada. Raramente, ocorrem movimentos transversais entre os fosfolipídos, ou seja, os fosfolipídos trocam de uma camada para outra.
Neste modelo, assim como os lípidos, também as proteínas apresentam grande mobilidade.
Existem dois grandes grupos de proteínas neste modelo: As proteínas periféricas ou extrínsecas e as proteínas integradas ou intrínsecas.
As proteínas periféricas ou extrínsecas são aquelas que se encontram à superfície e que se podem dissociar facilmente da membrana, por estarem associadas por ligações fracas às partes hidrofilícas de fosfolípidos ou proteínas integradas.
As proteínas integradas ou intrínsecas só se dissociam da membrana se ocorrer a disrupção da bicamada lipídica. Estas estão fortemente ligadas às regiões hidfrofóbicas dos fosfolipídos, podendo atravessar a membrana de um ao lado ao outro - quando tal acontece, têm o nome de proteínas transmembranares.
Na parte exterior da membrana celular, existem moléculas de glícidos associadas a proteínas - glicoproteínas - e por vezes associadas a lípidos - glicolípidos. Estas duas substâncias formam o glicoálix e são responsáveis pelo reconhecimento celular, ou seja, uma célula pode reconhecer outra célula semelhante a si através do glicoálix.
.jpg)
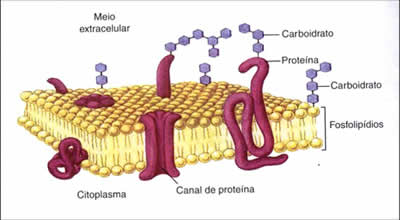
Transporte de substâncias na membrana plasmática
A membrana plasmática é uma estrutura que disponibiliza a passagem de várias substâncias em ambos os sentidos, ou seja, intracelular ou extracelular. A forma como a passagem é efetuada difere para cada substância. A percentagem de permeabilidade da membrana depende de cada substância, sendo em que alguns compostos consegue mesmo ser totalmente impermeável.
Existem dois tipos de transporte de substâncias na membrana plasmática:
Transporte Passivo: No transporte passivo não ocorre gasto de energia (ATP), pois as substâncias se deslocam do meio mais concentrado para o menos concentrado, ou seja, a favor do gradiente de concentração. O transporte passivo ocorre até que a concentração dentro e fora da célula sejam iguais. Existem três tipos de transporte passivo: osmose, difusão simples e difusão facilitada.
Transporte Ativo: No transporte Ativo ocorre gasto de energia (ATP), pois as substâncias se deslocam do meio com menor concentração para o meio mais concentrado, ou seja, contra o gradiente de concentração. Entre as substâncias que podem ativamente atravessar a membrana plasmática encontram-se os iões de sódio, potássio, ferro, hidrogénio, entre outros.
Transporte Passivo
Osmose
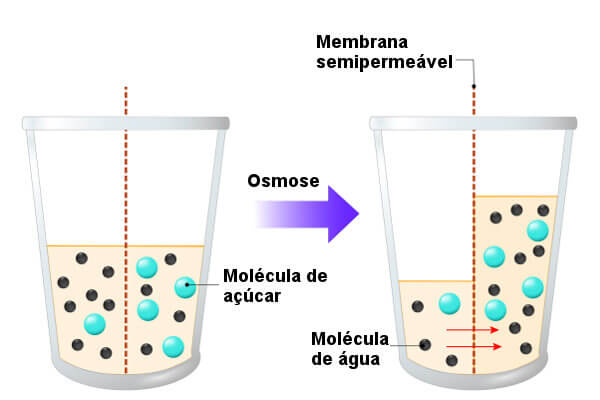
A osmose consiste na difusão das moléculas de água entre dois meios separados por uma membrana seletiva (permeável á água e pouco permeável ou até mesmo impermeável ao soluto) e é explicada pelas diferenças de concentração nos dois meios.
A osmose ocorre sempre do meio hipotónico (meio com menor concentração de soluto) para o meio hipertónico (meio com maior concentração de soluto). Quando estes dois meios atingem a mesma concentração de soluto (meio isotónico), o fluxo de água é igual nos dois sentidos.
Velocidade Osmótica
A velocidade osmótica é
proporcional à diferença de concentrações entre os dois meios. No início, como
a diferença entre concentrações é máxima, a velocidade osmótica também é
máxima. À medida que as concentrações dos meios se vão aproximando, a velocidade
vai diminuindo até as concentrações fiquem iguais e os meios se tornem
isotónicos.
Osmose em células animais

Se colocarmos uma célula animal num ambiente isotónico, a água flui na mesma proporção para dentro e para fora da célula.
Nessa situação, observamos que o volume da célula não se altera.

Numa solução hipotónica, observa-se um aumento da entrada de água na célula, nesse caso, a água aumenta o volume da célula rapidamente fazendo com que ocorra o seu rompimento (lise celular).

Em um ambiente hipertónico, observamos que a célula perde água para o ambiente, o que faz com que a célula possa morrer.
Percebemos, portanto, que uma célula sem parede celular sobrevive bem em ambientes isotónicos, porém o mesmo não acontece quando é submetida a condições hipertónicas ou hipotónicas.
Osmose em células vegetais

Quando colocamos uma célula vegetal em um ambiente hipotónico, a água entra por osmose nessa célula.
Entretanto, diferentemente da célula animal, ela não se
rompe, uma vez que a parede celular permite a entrada de água apenas até certo
ponto, passando depois desse período a exercer uma pressão contrária que impede
a entrada de água.

Quando a célula vegetal é colocada em um meio isotónico, não é possível observar uma tendência de entrada de água em grande quantidade na célula. Nessa situação, a célula fica flácida.

Em ambiente hipertónico, a célula perde água e murcha. A perda de água nessa célula faz com que a membrana plasmática fique solta em algumas regiões da parede celular. Dizemos que a célula sofre plasmólise.
Difusão Simples
A difusão simples é um tipo de transporte passivo de um soluto, onde não existe gasto de ATP, no qual as moléculas se movimentam do meio onde a sua concentração é mais elevada (meio hipertónico) para o meio onde a sua concentração é mais baixa (meio hipotónico), a favor do gradiente de concentração.
Quando é atingido um equilíbrio de concentrações, o movimento das moléculas não para, mas como o número de moléculas que atravessam a membrana num sentido é semelhante, não ocorre nenhuma alteração nas concentrações em qualquer um dos lados da membrana.

Devido às propriedades da bicamada fosfolipídica que forma a membrana celular, apenas moléculas
apolares lipossolúveis de pequeno tamanho (como a ureia, o oxigénio e o dióxido de carbono), podem
atravessar por difusão simples, sendo que moléculas polares terão que utilizar
canais proteicos através da membrana.
Difusão Facilitada
Na difusão facilitada, um elevado número de moléculas polares de dimensões consideráveis (glicose, aminoácidos ou algumas vitaminas) atravessam a membrana celular a favor do gradiente de concentração, também sem gasto de ATP, mas a uma velocidade superior do que o transporte por difusão simples.
Deve-se à existência de proteínas transportadoras (carregadoras) na membrana que ajudam na passagem destas moléculas. Estas proteínas são específicas para cada tipo de substância e são designadas de permeases.
A ligação da molécula a transportar à parte hidrofílica da permease faz com que esta modifique a sua forma, permitindo a passagem da molécula e regressando, em seguida, à sua forma inicial.
O processo de difusão facilitada efetua-se em três fases:
1.Ligação da molécula a transportar à permease;
2.Alteração conformacional da permease, que permite a passagem da molécula através da membrana e a sua separação da permease;
3.Regresso da permease à sua forma inicial.

O mecanismo responsável por limitar a velocidade da difusão facilitada baseia-se no facto de a substância transportada ligar-se a um sítio específico da permease. Dessa forma, quando todos esses sítios estiverem "ocupados", não adianta aumentar a concentração da substância a ser transportada. Quanto mais permeases (proteínas transportadoras) existirem, maior será a sua velocidade, mas se a concentração aumentar, a velocidade aumenta até chegar a um ponto em que estabiliza por não ser possível "inserir" mais permeases na membrana plasmática.
Transporte Ativo
Ao contrário no que acontece nos processos de transporte passivo, a célula também pode manter várias substâncias no seu interior. A membrana pode expulsar ou absorver alguma substância que esteja em excesso ou em falta, bombeando-a para dentro ou para fora da célula. Esta capacidade deve-se à permeabilidade seletiva da membrana celular e garante a manutenção das concentrações do meio intracelular, para que aí possam ocorrer as reações químicas vitais para a célula.
No transporte ativo, a célula gasta energia para mover uma substância contra o gradiente de concentração, ou seja, há transferência de substâncias de uma região de baixa concentração para uma região de alta concentração.
No transporte ativo, as mudanças de forma na proteína transportadora sucedem devido à energia que resulta da hidrólise de ATP, onde as proteínas transportadoras se comportam como enzimas, designando-se ATPases.

A
bomba de sódio e potássio é um exemplo de transporte ativo. Neste caso, a ATPase propaga a movimentação
do Na+ e do K+,
através da membrana, contra o gradiente de concentração. Através
da hidrólise do ATP em ADP e Pi,a
bomba de sódio e potássio,promove a saídade 3 iõe de na+ e a saída de 2
iões de k+, assegurando a diferença de concentrações nos meios intracelular e
extracelular.
Transporte de Partículas
Endocitose
As células possuem recursos que permitem o transporte, para o interior ou para o exterior da mesma, de macromoléculas , de partículas de maiores dimensões ou pequenas moléculas. O processo transporte deste tipo de material para o interior da célula por invaginação da membrana plasmática designa-se endocitose.
Existem 3 tipos de endocitose: fagocitose, a pinocitose e a endocitose mediada por recetor.
Fagocitose
Na fagocitose, a membrana celular engloba partículas de garndes dimensões ou até mesmo células inteiras.
Ocorre quando a célula emite prolongamentos que englobam a partícula (pseudópedes) formando assim vesículas fagocíticas ou fagossomas (com um diâmetro superior a 250 nm). As vesículas formadas fundem-se, geralmente, com lisossomas (vesículas que contêm enzimas digestivas), originando vacúolos digestivos, onde se dá a digestão das substâncias fagocitadas.

Pinocitose
É um processo semelhante ao da fagocitose, em que e as substâncias que entram na célula são substâncias dissolvidas ou fluidos. Por este motivo, as vesículas são de menores dimensões. Ocorre por exemplo no epitélio intestinal e acontece de forma quase contínua na maioria das células eucarióticas através de pequenas vesículas (com um diâmetro inferior a 150 nm ), contendo fluidos e solutos.

Endocitose mediada por recetor
A endocitose mediada por um recetor ocorre quando as macromoléculas entram na célula, ligadas à membrana das vesículas de endocitose.

Exocitose
Processo inverso à endocitose, onde as células são libertadas para o meio extracelular. Neste processo, as vesículas de secreção fundem-se com a membrana plasmática, libertando o seu conteúdo para o meio extracelular.
Este processo é essencial pois é assim que a célula se livra de resíduos de digestão intracelular, mas também pode fazer parte do processo digestivo de seres pluricelulares. É por este processo que são segregadas as enzimas digestivas do pâncreas.
